
Coincidences, mysteries, personal lessons, and strange experiences of the most diverse character, as significant and soul-searching as the Mexican earthquake experience, continued with a very suspicious frequency. I have previously mentioned my ability to bear up under the influence of any drug or shamanic potion, and I sometimes found this ability of even greater value in dealing with the people and day-to-day events playing a part in the unfolding of my work. The idea that the shaman himself must deal with forces and destinies on a far more fundamental level than the members of his tribe is, of course, the very spirit and tradition of shamanism. If powerful psychedelic experiences were to benefit members of the tribe, the shaman must have already explored the same territory, and gone well beyond to the very limits of his abilities. In the present case where the shaman not only administers the powerful medicines, but actually creates them with the tools of modern science, his knowledge and intentions are perhaps even more important.
Thus, in the case of what turned out in some ways to be a rather successful collaboration between myself and a trio of avowedly enthusiastic partners, the passage of time and the achievement of some degree of success seemed to distort the relations between us in a way very reminiscent of a kind of vicious parody of the psychedelic experience itself. Meetings and discussions of plans and goals at first were nothing less than inspiring. Between us we had the connections and ability to obtain the necessary raw materials, manufacture a high-quality product, and distribute it in such a way that it would reach the right people.
We certainly did not advocate or intend any sort of mass distribution of psychedelics resulting in their misuse or ignorant use, which would only call attention to a situation easily besmirched by adverse publicity. Sensational, fear-mongering publicity had already made it much more difficult to properly initiate or introduce a newcomer to the psychedelic experience. There was certainly a case to be made that many negative experiences were a direct result of the adverse publicity itself, for in the early, pre-publicity days of psychedelic research, "bad trips" were a rarity, even among alcoholics and psychiatric patients. With normal research volunteers, the statistics noted by the many researchers indicated the astonishing safety of psychedelic drugs, not risk. Now the publicity made it seem that, although an Aldous Huxley or Alan Watts might get through a psychedelic experience unscathed, the normal member of society should realize that the potential risk was overwhelming. And this kind of official attitude weighed heavily on the person interested to undergo an experience: even if such a view could in principle be seen for the hysterical, prohibitionist, puritanical mindset that it in reality was, the slightest lingering doubt had the possibility to poison a person's trust in himself and his shaman.
It is perhaps easier to see demons where there are none to be found. And if demons are encountered, the idea that they are exterior, real, and caused by something other than oneself is certainly not conducive to understanding their meaning. As the Habit Routine Suspension model demonstrates, the psychedelic experience is no roll of science-fiction film projected upon consciousness from the outside. If demons are encountered, they are in reality our everyday friends seen without the normal range of categorizations which render them ordinary and insignificant.
The demons which appeared to disrupt our successful enterprise were therefore more real than those of the imagination. I can only suspect a hidden agenda on the part of my co-workers, but the nature of their intentions at the end was so at odds with the honest enthusiasm of the beginning that I suspect that the hidden motivations must have developed over time. From the original enthusiasm there seemed finally to have evolved a plan to demonstrate once and for all that the psychedelic experience was, in fact, illegitimate, or at least unnecessary, its insights illusory, its history merely primitive self-delusion. To arrive at the establishment reactionary position through first having professed a more universal and enlightened view seemed to me altogether impossible. This demon was real. And what was I going to do about it?
The final scene was quite surreal. The night before, one of my partners presented me with a five-hundred dollar banknote, subtly making sure that I noticed the portrait on the face of the bill, that of the "assassinated leader", William McKinley, twenty-fifth president of the United States. At most, I felt only the slightest premonition of the events the next day would bring, for which we were planning a day of skiing, and restaurant to follow. Whether the banknote, and perhaps other similar but unrecognized intimations were apparent I do not remember, but in the morning I decided to take a small dose of LSD, to appreciate a day in the high snowy mountains from a different perspective. In retrospect, I find that I instinctively tended to undergo a psychedelic experience at points in my life where a certain crossroads was about to be reached, when perhaps only vaguely realized intimations of important changes to come had appeared. Without fail, the experience would precipitate whatever it was that was pending, and ignite the insight and decision that would usher in a new direction.
My friends were newcomers to skiing, and I had promised to help them get their sea-legs, so to speak. Upon arriving, I sent them off to the rental shop to secure their equipment, and I took a test-run on the nearest slope. It was exhilarating! Although there was quite a crowd that day, the scene was magnificent. Arriving at base, my friends had not yet reappeared, so I continued on up again to the irresistible heights. The next time down the crowd was getting thick enough to make locating my friends problematic, and in my present state of exhilaration I couldn't begin to devise a sure-fire method for finding them. I had to assume that, although our original plan seemed to be going astray, they would nevertheless have suited up and found some incidental help with their first few glides on skis. After a few more runs I began to feel, in contrast to my exhilaration, a gnawing guilt that I had ignored my friends, even if through a concurrence of events which I could not have foreseen. The LSD experience, despite the very modest dose that I had taken, was attaining a peak more spectacular (in more ways than one) than those amongst which I was gliding.
Suddenly my friends appeared, and I knew that my neglect had ruined their day. But the intensity of my feelings, and their reaction as well, seemed all out of proportion to the reality of the situation. There was far more significance afoot than a simple evaluation of the day's events would warrant. My apologies, and their subdued yet somehow exaggerated insistence that it didn't really matter, were a mere facade for the emotion that was all too evident in just a glance. Somehow aspects of the entire relationship in our joint involvement in the psychedelic project were metaphorically represented here, and they were threatening a tumultuous final act upon the stage whose players had to date seemed so reassuringly without calculation or subterfuge.
The situation while driving back continued to intensify. I began to see through the cracks in reality, the physical manifestations of which became more and more a flimsy veil affording less and less protection against immediate dissolution into the hidden dimensions beyond. If my friends were worried about my driving on the slippery roads, I think that they too must have sensed, more importantly, that loyalties and intentions concerning our collective enterprise was the real issue that was coming to a head. At the restaurant, under the influence of good food and wine and associated conversation, the electricity subsided somewhat, and it seemed that things might be patched up with a resulting return to a situation that we all now realized had more of deception than honesty. If we could return to business as usual, we all now knew that on a more fundamental level, all was changed. This is not, and never has been, a situation from which effective shamanism can work its benefits to the full.
Later at their house, in front of a warm log-fire, I continued to ponder on this last point. So far, absolutely nothing explicit had actually been discussed concerning the impending crisis that we each knew to be imminent. An outsider could not have detected that anything more than the superficial was transpiring, yet in each others' presence, a mere glance was like a trumpet fanfare announcing major discoveries about a hidden side of our co-involvement. It must have been past midnight when in walked partner number three, whom I had seen only a few times. He lived quite far away, and I was surprised to see him at this late hour, but the others apparently had expected his arrival. After some lengthy and private discussion in the kitchen, they reappeared, and, very tentatively at first, began to suggest to me a most surprising plan.
I had recently been working on some advanced techniques for making our product in a more purified form, and had been meditating about my experiments probably as a way to suspend thinking about the more serious implications of the day's events. At my partners' first mention of LSD I immediately launched into an attempted discussion of my recent experiments, perhaps also hoping to repair the situation: At every stage of my work it was of primary importance to me to be able to continue, even if this involved having people associated with the project who neither knew nor cared about the larger implications. If a partner was interested in nothing but the cash-flow, for example, I tried to cope with the deficit temporarily until something better presented itself.
But something was seriously wrong here. One of my partners was now saying to me that he didn't like the effect of LSD, it seemed dirty to him, made him sick rather than giving him inspiration. Despite the day's events and the suspicions that had been aroused I was quite shocked, and speechless. Gradually the hidden agenda was presented: what I was to do, was to prepare a large batch of placebos. Make up what I would say was a batch of the purest LSD that had ever been made, give them away free to all my contacts for distribution, and pass along the message that everyone was to wait until the vernal equinox, then take the new psychedelic together. The trick was, they would be entirely inactive, blank doses with nothing but an implied message from... Yes, from whom? Who was the author of this message? Was this idea a brainstorm of my partners, suddenly having seen a vision that drugs were the tools of the devil? The apparent honesty of our relationship so far in contrast to the way that the plan was presented, and the things that were said that night, including the implied threats (the McKinley banknote was burning in my pocket), and also the implied promises of new and greater projects and responsibilities for me if I could only see the wisdom of this plan, made it plain to me that the tiger I had by the tail was rather larger than I had heretofore suspected.
Well, the activation of my consciousness by the dose I had taken, and these astounding changes of identity made manifest that day, made the indoctrination as effective as a lengthy torture and brainwash session in North Korea. I quite forced myself to believe the idea had some merit, and for the next few days, more than half-heartedly prepared a large batch of chocolate covered placebos in line with the master plan. The chocolate coating was to prevent anyone examining the underlying substrate and possibly suspecting that there was less than meets the eye about this plan for a glorious first-day-of-spring celebration.
I carried through with the lie to the bitter end, I don't think there were any suspicions, and I myself boarded a train for the Grand Canyon to be (1)out of reach when the chocolate hit the fans and (2)in a nice spot in case of the highly unlikely event that the Millennium was actually going to begin that day. It didn't, and the feedback from the experiment was less than respectful. My former partners disappeared from the scene to cope with more petty problems and new coincidences materialized out of nowhere to enable the next stage of my ongoing quest.
There is an old Negro Spiritual that was in the 1940's made into a "Jazz Soundie", the equivalent of today's rock video as seen on MTV. I have a wonderful 3-minute version of "Dem Bones" by the Delta Rhythm Boys, sung in close four-part harmony; it starts with the refrain,
Ezekiel connected dem...DRY BONES,
Referring of course to Chapter 37 of the Old Testament Book of Ezekiel in which the prophet is commanded to "Prophesy upon these bones: and say unto them, O ye dry bones, hear the word of the Lord." Whereupon, as in the refrain of the song,
Oh, de toe bone connected to de...FOOT BONE,
De foot bone connected to de...HEEL BONE,
De heel bone connected to de...ANKLE BONE,
And so on up to "de HEAD BONE". I hadn't watched my collection of "Soundies" in quite some time, but humming in the shower late one night, meditating on my recent neurological studies and lack of satisfaction of their power to explain processes of mind except in a very rudimentary manner, the tune suddenly popped into my headbone. It occurred to me that current neuromechanical models of how the collection of parts in the brain is supposed to produce mind, behavior, thinking and perception, was very much a Dry Bones Model of Neuromechanics. (Substituting brain part names into the song quite ruins the rhythm however: de caudate nucleus connected to de...CEREBELLUM, de locus coeruleus connected to de...ANTERIOR CINGULATE GYRUS, you get the point.)
The textbooks carefully told the student what was connected to what, and suggested sometimes that a particular part "was thought to be involved in" some function or other. And to my further dissatisfaction, slowly compiling again and again my own map of brain connections, my studies had revealed that practically every brain part was connected to every other brain part: each brain map I plotted quite soon became filled up with arrows, and quite useless. It seemed that mainstream neuroscience, in spite of its great and rapidly increasing wealth of assuredly precise data, had little to say about the functions of all these parts on a systems level; understanding how to get to psychology from neurology, how to get mind out of brain, was still at the level of expecting dry bones to suddenly, as the song's second verse goes, after "dem bones" have been connected,
Dem bones, dem bones gonna'...WALK AROUN'...
The neuroscientists, without such divine intervention as Ezekiel had access to, simply were not able to show decisively how connections of the nervous system could produce even simple psychology, much less consciousness itself. My own efforts to suggest brain systems and connections that might allow the operation of the habit routine search cognitive process would therefore be no less tentative and imprecise, certainly due for major revisions as new evidence was discovered.
In the case of memory, so fundamentally important to everyday life and to the operation of my proposed habit routine system, it is therefore still very much a matter of debate as to how information is stored in or associated with the properties of the neurons in the central nervous system. The "data" representing the various types of memory, including habit routines, must in some sense be "in the brain" and by extrapolation must somehow be associated with the properties of neurons and their connections, but the wide diversity between current theoretical viewpoints illustrates more our lack of precise knowledge than an emerging paradigm. Perhaps the most popular model of information storage at present is that of the adjustment and long-term maintenance of the strengths of the connections between neurons. This is supposed to occur by the modification of the neuron's synapses, the junctions by which neurons communicate using chemical neurotransmitters.
This "neural network" or "synaptic weight" model lends itself to ready simulation on a computer and is probably on one end of a scale ranging from extreme reductionism, to another extreme which reductionists usually brand as mysticism. An example would be the ideas about memory advanced by Rupert Sheldrake, that the actual memory information is resident in some quasi-independent memory field which may allow such phenomena as telepathy, apparent reincarnation and so forth, the physical brain being merely the sender-receiver of memory to the common field by a process of morphic resonance. (1) In looking at the broad range of sometimes very enigmatic properties of memory, and the admitted uncertainty and disparity of views by the experts, no model should be routinely dismissed if it has any explanatory power whatsoever.
The synaptic-weight model probably owes its status to the power and success of computer science, the enormous investments and hence large number of talented scientists working therein providing a momentum of opinion which may unduly limit the credibility of other well-constructed but competing models. It cannot be denied that very impressive mathematical models and computer simulations of "intelligence" have been demonstrated, and shown to have similarities to the ways in which we believe human intelligence to operate. It has also been well demonstrated that synapses between neurons (as well as the size and connectivity of the neurons themselves) can and do change their characteristics for varying lengths of time under the influence of learning processes. The power and sophistication of neural network models (see for example descriptions of the operation of the Hopfield Network as a mechanism for the storage/retrieval of information) (2) are certainly impressive. The fundamental question which remains to be resolved, however, is whether the brain works anything like a computer. There is currently raging a most entertaining debate on this subject, some of the most powerful intellects of our time are joining the battle, (see, among others, works by Churchland, Claxton, Crick, Dennett, Edelman, Flanagan, Fodor, Freeman, Gazzaniga, Hofstadter, Humphrey, McGinn, Penrose, Searle, Tallis, Tulving, the list is so long there is no recourse but to leave out mention of many more. Some titles are listed in the Bibliography).
Steven Rose, himself a veteran of many years of research attempting to identify brain sites involved with storage of engrams or memory traces, has recently summarized criticism of the computer-like, information-processing model of brain operation. (3) Particularly of interest, as Rose points out, is a critique by the neurophysiologist Walter Freeman, for it is solidly based on research findings: Freeman insists that, although changes to various individual neurons can be observed to happen as a result of learning, the "information" that is learned does not subsequently exist as "bits of data" recorded in these neurons (the computer paradigm). Rather, the memories of the learning exist as fluctuating dynamic patterns of electrical activity generated by the entire brain. As Rose puts it,
[The] experiments say that it, the engram, is not confined to a single brain region. But I want to go further than this, and to argue that in an important sense the memory is not confined to a small set of neurons at all, but has to be understood as a property of the entire brain, even the entire organism. (4)
An intriguing suggestion has been that the storage of information in the brain is analogous to the storage of information in a hologram, a model which was first developed by the neurosurgeon Karl Pribram in the 1960's. (5) The idea that brain activity might be non-local, a process of the generation of distributed waves of activity which interact to form interference patterns, had been suggested in 1942 by Karl Lashley, whose pioneering research on the brain had inspired Pribram during their collaboration at Yerkes Laboratories. Freeman's experimental findings, as well as the observations of Steven Rose above, are exactly what one would expect from a brain operating on holographic principles. The view of memories existing as "fluctuating dynamic patterns of electrical activity generated by the entire brain", could not embody holographic principles more strongly.
Although criticism of the so-called holographic paradigm has come from many directions, the parallels between the mathematical and physical facts of optical holography and many known properties of memory and the brain suggest that further research will tend to support rather than discredit the model. In a recent paper Pribram elaborates on some of these parallels:
The following properties of holograms are important for brain function: (1) the distribution and parallel content-addressable processing of information — a characteristic that can account for the failure of brain lesions to eradicate any specific memory trace (or engram); (2) the tremendous storage capacity of the holographic domain and the ease with which information can be retrieved (the entire contents of the Library of Congress can currently be stored on holofische, or microfilm recorded in holographic form, taking up no more space than is contained in an attache case); (3) the capacity for associative recall that is inherent in the parallel distributed processing of holograms because of the coupling of separate inputs; and (4) the provision by this coupling of a powerful technique for correlating (cross-correlations and autocorrelations are accomplished almost instantaneously). (6)
It is a curious fact of the history of neuroscience that Pribram's work during the 1960's and 1970's was almost unanimously rejected by the mainstream: he was for two decades practically the only proponent of the theory of distributed representation of memory (distributed coding) in the brain. At the time, nearly all theoretical work leaned strongly toward the assumption of detector-cell coding, (7) a model strictly analogous to computer, bit-storage processes, with a precise location in the machine being responsible for the storage of a uniquely defined unit of information. Today the mainstream neuroscientists have taken the opposite tack: distributed processing and storage is their byword, yet Pribram's holonomic theory is still widely rejected, even though in an important sense it is part of the foundation of recent theory. Francis Crick's assessment is typical:
This analogy between the brain and a hologram has often been enthusiastically embraced by those who know rather little about either subject. It is almost certainly unrewarding, for two reasons. A detailed mathematical analysis has shown that neural networks and holograms are mathematically distinct. More to the point, although [artificial] neural networks are built from units that have some resemblance to real neurons, there is no trace in the brain of the apparatus or processes required for holograms. (8)
To state that neurosurgeon Pribram and his recent mathematician colleagues "know rather little about either subject" is a bit stiff, and the observation that there don't seem to be any lasers or such holographic equipment in the brain itself demonstrates ignorance of the two subjects. Pribram counters such criticism with the warning, "It is important to realize that holography is a mathematical invention and that its realization in optical systems...is only one product of this branch of mathematics." In other words, optical holography is merely a special case of a wider and more fundamental process for the encoding and reconstitution of information. For this reason it was suggested that the term holonomic (9) be used in reference to brain/mind properties, a term I shall adopt here. The term holonomic as used in physics, also indicates that the process is not a static, frozen-in-time representation as is a hologram, but a dynamic, continuous one.
Further rebuttal, including a short history of the criticism of the holonomic theory can be found in a recent paper by Pribram and his colleagues. (10) The most recent and complete exposition of holonomic theory is presented in a book covering the 1986 John M. MacEachran Memorial Lecture, (11) delivered by Pribram. The book demonstrates that the holonomic theory has progressed both conceptually and mathematically far beyond its introductory position of the 1960's; it is the original and necessarily simplistic exposition of the theory which is still attacked by the critics of today who remark (along with other trivial complaints) that "there don't seem to be any lasers in the brain".
Other criticism of the holonomic model of memory storage and retrieval has been derived from the perceived contradiction with the accepted dogma that regulation of synaptic weight must certainly be the only possible storage mechanism in the brain. Synaptic weight distributions are quite amenable to neural network, viz. computer-like models of brain/mind function, but it was very difficult to see how synapses could function as a storage medium for the interference patterns implied by holonomic models. Yet the idea of learning and memory storage by neural networks is itself by no means universally accepted. Concerning the much-touted ability of computer analogues of neural networks to learn, Jerry Fodor remarked: "Much has been made of this, but, in fact, it's a tautology, not a breakthrough." (12)
A very promising idea which would obviate some of these objections to the holonomic model has been the recent suggestion that the site for storage of information is not the synapse per se, but associated with the microtubules making up the cytoskeletal structure of the neuron itself. (13) (The synapse is known to be in indirect yet intimate contact with the microtubule structure of the neuron.) This model has the immediate advantage, as Hameroff points out, of increasing the potential effective complexity and storage capacity of the brain by about seven orders of magnitude, (14) but there are many other attractive aspects to the model as well.
It would seem that the synapses are concerned primarily with the large-scale systems of the brain, they are the functional units enabling coordination and signal transfer between distinctly separate, sometimes distant locations. For example, they enable the experimentally-demonstrated phenomena of the sequential processing of sensory information from primary sensory cortex through several levels of associational cortex. Another example would be a neuron in a brain-stem nucleus such as the locus coeruleus: it may project the entire distance to an area of the cortex before synapsing on another neuron. As a first consideration, the synapses thus seem involved with the switching and transfer of information (analogous perhaps to the transistors or switches in the central processor of a computer). To assign them the double duty of also being the storage site for the information which they manipulate seems merely a default position (the synaptic-weight model). There is more logic to the idea that the information that is manipulated via the synaptic connections has been accessed in structures or locations independent from the synapses, otherwise the duty of the synapse in one function would seem to necessarily limit its capacity for the second function.
Having looked into the neural network models of brain function, I realize how easy it would be to reject the foregoing argument if one is convinced of the validity of such a model. But it is my own view that, although neural networks seem to simulate brain/mind operation in some trivial respects, the "connectionist" paradigm as it is called is as primitive and misleading a model of the human brain/mind as is the planetary model of the atom (discussed in chapter 2). From this viewpoint, neural networks may be useful for modeling "intelligent" processes up to the level, say, of creating robots for assembly-lines or for playing chess, but to extrapolate from there to say that by adding a few more bells and whistles, or even many additional layers of mechanical or electronic complexity to these beasts, we will then have duplicated the human mind in a machine... Well, to me and many others that is sheer folly. I am all for letting the connectionists connect, and the AI (Artificial Intelligence) crowd continue their quest to reproduce themselves asexually, but they do tend to pummel the less-well financed competition with epithets worthy of the flat-earth advocates of a bygone age. Hence the present riposte.
The combination of the holonomic storage/retrieval model with the hypothesis of micro-tubule based storage sites thus seems even more promising: Hameroff's paper even suggests that the microtubules of neurons might provide a location where coherent photons would operate (viz., the kind of light necessary for optical holography). In constructing my own model of brain operation, I have tried not to depend irrevocably on any particular view now contending, including the more conservative ones. Yet due to the wide disagreement between the positions of the several groups of theorists, I have felt justified to freely use some of the more speculative suggestions such as holonomic operation and storage, if only as temporary visual, heuristic devices. Since my intention is merely to suggest a possible mechanism for the operation of the habit routine system, (and at the present stage of neuroscience, it would be unrealistic to hope for more), my overall theory of psychedelic experience would not be damaged by further research indicating the brain had many further tricks up its sleeve. The holonomic, microtubule hypothesis, although it may well be found to describe the actual operation of the nervous system, will for my purposes merely provide a convenient conceptual model facilitating the understanding of the operation of the brain systems responsible for habit routine search and suspension and other proposed operations involved with psychedelic experience. I thought it valuable, however, to introduce the holonomic model on a level which would show that it was far more than "New Age Techno-Babble", an implied if not explicit critical view now prevalent.
Figure 2 illustrates the location of just a few of the major brain-parts for the benefit of the general reader. But in the interest of brevity and concision, I will not attempt to diagram or explain entry-level details of the model I shall now present, either in the case of the properties of neurons, or the general views concerning the functions of the various brain components taking part in my own theoretical view. For the layman, the Glossary will be of some help, but a survey of a textbook such as Principles of Neural Science (Kandel, Schwartz & Jessell), would be indispensable for evaluating the proposed model in detail, and comparing it to standard textbook views representing current paradigms.
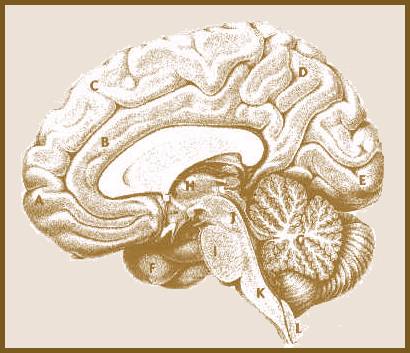
FIGURE 2. The Right Half of the Human Brain.
Viewed from a vertical plane bisecting the brain into its left and right hemispheres. A through F: Areas of the cortex. A: Prefrontal area. B: Anterior Cingulate Gyrus. C: Frontal Lobe. D: Parietal Lobe. E: Occipital Lobe (primary visual cortex). F: Temporal Lobe: most of this area of the cortex is on the outside of the far side of the brain, hidden in this view. G: Cerebellum. H: Thalamus. Encircling the Thalamus are the Hippocampus, the Amygdala, the Basal Ganglia and other parts of the limbic system. I: Pons. J: Midbrain, area in which are found the various brain stem nuclei including the Raphe Nuclei, the Locus Coeruleus, the Substantia Nigra, etc. K: Medulla. L: Spinal Cord.
(Modified from Nieuwenhuys, et. al., The Human Central Nervous System).
I have mentioned that most brain areas are connected to most other brain areas, the multiplicity of connections between parts becoming obvious and soon quite bewildering when one starts examining the various brain "maps" in Nieuwenhuys' book, for example (the source of the diagram in figure 2). In addition, it is found that nearly all of the connections are two-way, the connections between the thalamus and the primary sensory areas of the cortex, for example, being reciprocated by nerve pathways (backprojections) in the opposite direction. It is problematic therefore to construct a diagram for the supposed "flow of information" in the brain: if the retina of the eye, for example, sends its "visual data" to the thalamus (which has been likened to a relay-station for sensory data), and then the thalamus sends this signal to the primary visual cortex for the first stages in the processing of visual information, why should the primary visual cortex send a nerve pathway directly back to the same area of the thalamus from which it has just received the data? The backprojection is not insignificant: recent findings indicate in the case of vision that there are ten times as many nerve fibers in the "backwards" direction as in the direction in which "information" is supposed to flow! The size and importance of the various interconnections (15) seems to indicate that the actual sensory "information", the environmental data ENV in figure 1, constitutes only a minor part of what is being "processed" in the brain! Current computer "data-flow" models of brain operation cannot explain the facts of the existence and relative importance of backprojections, as even the best workers in the field will admit.
But this is exactly what one would expect for a brain architecture that operated according to the habit routine model of cognitive function. The environmental data merely provides cues for the elaboration of the far more complex informational entity of which we become aware, the habit routine complex. It is far more complex, because it is generated from the entirety of previous experience stored in memory, whereas the actual environmental data is quite limited in scope not least by the limits of the sensory organs themselves. In size and complexity, the nerve pathways carrying the primary sensory information to the cortex are among the less important connections of the brain. To be sure, the environmental input is necessary: when you shut your eyes, vision promptly ceases. But this effect itself may be heavily dependent upon the operation of habit routines. We are all absolutely and automatically certain that vision must cease when we close our eyes, and would be profoundly confused, (if not shocked into psychosis!), were it to be otherwise, even for a few seconds.
There is a computer analogy that could be made in the attempt to account for the curious facts of neural reciprocating connectivity, that the signaled area must send back a return signal indicating that it has received information, like two modems do as they talk to each other. But this would not explain why return pathways are so much larger. I would propose that something much more interesting is taking place. In the case of the nerve pathways from the thalamus to the primary sensory cortex areas (and back again) for the various sensory modalities, I believe it is useful to hypothesize that a reverberation is being established with the two-way signaling, and that this reverberation is a dynamic informational entity having holonomic properties.
The thalamo-cortical reciprocating nerve connections set up for each sensory domain a dynamic reverberating holoprojection of information, which is constantly updated and modified with the newly arriving signals from the sensory organs. It would require a much higher density of nerve pathways to set up and maintain such reverberation than to feed in the flux of newly arriving ENV data, thus explaining the relative importance of the brain connections between the sensory receptors, the thalamus, and the areas of the sensory cortex. The suggestion of similarity to the projection of a holographic image is intentional, for I believe that, not only are the mathematical principles which predict and describe optical holography applicable to memory storage, but also to the ongoing operation of many of the systems of the brain. In comparison to optical holography, it can also be maintained that the relation between a given unitary nerve signal (the electrical action potential of a neuron) and the overall holoprojection to which it contributes, is analogous to the relation between the unitary nature of one grain of photoemulsion making up a hologram (the "photograph" of the interference patterns produced in optical holography), and the resulting projected holographic image. The single grain of emulsion on the photographic plate may only be either "on or off" like the neuron, yet it theoretically represents the entire holographic projection, (16) albeit with a resolution of zero. It is the same for a single action potential: it represents the entire holoprojection, but with zero resolution.
Someone familiar with holography would certainly ask, but how and where are interference patterns produced, certainly any holonomic process implies their existence, for it implies the interference of two or more signals? Dropping one pebble into a still pond produces concentric waves, but dropping two pebbles produces an interference pattern between the two sets of waves. So it may be with the nerve signals of the brain. It is well known that neurons in their various nerve pathways have a background rate of firing which, for all intents and purposes, seems to be merely random noise. Here is pebble number one. Pebble number two (in the case of the primary sensory holoprojections), is the impinging signal from ENV (of figure 1), the signal coming from the sensory receptors.
Thus the resulting holoprojection is the product of a dynamic interference pattern resulting from at least two distinct signals, and is amenable to expression as mathematical transform coefficients analogous to the mathematical operations which describe optical holography. In the nerve pathways maintaining a primary sensory holoprojection, the microtubules of these neurons record and dynamically maintain the transform coefficients which represent the information necessary for the neuron firings to maintain the reverberation. The coefficients are constantly updated with the sensory signal from the environment, which also exists as a transform of the interference patterns actually received by the sensory receptors. Thus there are two sets of coefficients representing the two signals, together they contain the information necessary to maintain the dynamic holoprojection in time. It will be seen that even the background firing of the neurons, the resident signal, is not merely random noise, for it is generated from the coefficients resident in the microtubules and represents the holoprojection in temporal cross-section. The constant arrival of the ENV signal produces the dynamic aspect of the primary holoprojection.(note)
But the combination of signals to produce interference patterns does not end with the primary sensory holoprojections, for as I shall explain below, holoprojections themselves combine and overlap, they become superimposed under the guidance of certain brain components so as to produce further interference patterns and thus further composite holoprojections. It can be seen that the "processing of information" in the brain is therefore accomplished using entire simultaneous fields of bound "data" from several, or even the entirety of all ongoing processes. The hypothesis of such a process conflicts radically with the computer, neural network model of the brain in which the serial processing (in parallel pathways) of discrete bits of information is the proposed mechanism. If experimental results begin to confirm the holoprojection model of brain operation, they will be a significant argument against the pursuit of strong Artificial Intelligence as it is presently conceived. Let us see how the fields of information I have called holoprojections might function in stages of brain operation beyond the primary sensory realm. First let us take a closer look at the thalamus, which plays so central a role in generating the primary sensory holoprojections which are the data fields upon which all further brain activity is based.
The thalamus itself is composed of many different nuclei, widely connected to other brain areas and interconnected as well. Thus in discussing the various functions of the thalamus, it must be kept in mind the great diversity of independent yet interrelated parts and functions comprising this centrally-important component of the brain. In the case of vision, after the signals have passed from the retinae through the optic chiasm which combines and distributes the visual signals from both eyes to the left and right hemispheres of the brain (the thalamus also is divided bilaterally), the visual signal that is to be used for updating the primary visual holoprojection enters a thalamic nucleus called the lateral geniculate nucleus, or LGN. The LGN then signals the first stage of the part of the cortex involved with vision, the primary visual cortex, and it is from this area that we note the very important nerve pathways which return directly to the LGN. It is merely a convenience to say, for example, the signal is "passed" from here to there, and "then" passed...etc., for we must remember that all these processes are dynamic, continuous, and as I have proposed, reverberating and holonomic.
The function of the LGN of the thalamus thus appears to be as a "driver" for the reverberation between the LGN and the first stage of the visual cortex. As I have suggested, this reverberation may be thought of as a holoprojection which has at least two functions. Firstly, this informational entity is the first stage in the generation of (the visual aspect of) the habit routine complex, it contains the information which will activate from the frames of memory (stored in distributed manner in the same visual areas of cortex), the actual information which makes up the habit routine presented to thinking2 processes. Secondly, since this holoprojection carries, at least potentially, the original or "genuine" visual information, it will be used under certain circumstances to generate aspects of the visual scene that are detected as significant and to which the attention is directed. This will occur by a comparison or superimposition of the primary visual holoprojection with another holoprojection set up by the habit routine search system. (As explained below, significance detection may use the primary sensory holoprojections, or the memory data activated by these holoprojections instead).
An additional function of the primary visual holoprojection results in the phenomenon of iconic memory, the very short term visual memory trace that has been experimentally demonstrated. (17) An informational fragment of the primary visual holoprojection, since the entire holoprojection is being constantly updated with new visual data, would be expected to have a very short "half-life" comparable to the 200 to 500 millisecond iconic memory (up to three or four seconds under certain experimental conditions). Experiments with iconic memory have shown that it persists for greater periods when preceded and/or followed by a simple dark field containing little or no new visual information. Likewise, it may be interfered with by the process of masking in which a bright field of view, or interfering patterns are shown. This is exactly what would be expected for the visual holoprojection model. A paucity of new visual data arriving to update the holoprojection would allow the reverberation to persist "as is" for a greater length of time. Data intentionally designed to confuse or interfere with the iconic memory would have the effect of overwriting the relevant aspects of the holoprojection (the transform coefficients stored in the microtubules of these neurons).
The extraction of the iconic memory must occur by other systems involved with thinking2 processes, again probably by the comparison or superimposition of holoprojections. There is more than speculation in the idea that such projections might be superimposed and compared to show similarities and differences, for the same kind of process can be carried out with optical holograms (in practice, I am informed, there are technical difficulties to overcome, but no theoretical restrictions). Two slightly different optical holograms, for example, could at least theoretically be projected so that they cancel each other except for the aspects in which they differ: only the differences would appear in the projected image. Likewise, two radically differing holograms having just a few identical features could be projected to emphasize their common features. In the brain, the process might be as simple as the addition and subtraction of the transform coefficients stored in the microtubules of the contributing neural systems. This would produce another set of coefficients representing the superimposition.
At the same time that the primary visual holoprojection is being generated, of course, all the other sensory systems are generating their own holoprojections, by similar mechanisms involving sensory receptors, the thalamic relay nuclei concerned with those senses, and areas of the cortex. Thus for hearing we get an audio holoprojection from which can be extracted echoic memory (analogous to iconic memory). The audio holoprojection is used to activate the audio domain of the frames of memory going into the generation of the habit routine complex. Tactile sensations and proprioception likewise produce their reverberations, and so forth.
The binding and superimposition of all the primary sensory holoprojections is accomplished by a scanning mechanism only recently detected by neurological experiments. (18) This scanning operation is also carried out by a nucleus of the thalamus, the intralaminar nucleus. Now what is scanned is not the actual primary holoprojections themselves, but the memory information which they activate in the various regions of the cortex. This activation occurs as the holoprojection signal transits through the pyramidal cells of the several layers of the cortex taking part in the reverberation. The sum total of all ongoing sensory holoprojections, impinging on the memory data distributed in the same areas of the cortex that take part in the set up of the various holoprojections, activates this memory data such that the intralaminar nucleus, acting again as a driver for the generation of a holoprojection, creates the habit routine complex holoprojection. The memory data in this process is thus analogous to the role of primary sensory data in the generation of the primary sensory holoprojection. Remember that we are hypothesizing that this memory data is associated with the microtubules of the neurons in the cortex and not their synapses, so while the neurons and their synapses in the circuit between the thalamic relay nuclei and the cortex are maintaining the primary holoprojection using the signals from the sensory receptors, the holoprojection itself, as an interference pattern, is resonating with the stored interference patterns in memory resident in the microtubules.
The habit routine complex holoprojection is the bound informational entity presented to thinking2 processes, having the various properties already described in chapter 3. The memory information used to generate the habit routine is, as just proposed above, analogous to the original sensory information: the sensory signals are used to set up the primary sensory holoprojections, and the activated signals from memory are used to set up the habit routine holoprojection. In each case we have a thalamic nucleus acting as the driver for the process, using an input of transform coefficients to produce the holoprojection. But the intralaminar nucleus scan which sets up the habit routine is deriving its input from the entire cortex, not just the primary sensory areas. The habit routine complex is much more than just primary sensory information taken from memory, for remember that a habit routine contains pre-programmed associations with ideas, with habits of thinking, and it also contains recommended actions (including not only physical responses but thoughts, opinions, implied value judgments, etc.), that represent the sum total of ways in which similar situations were dealt with or reacted to in the past.
The higher domains of the cortex itself are using the primary sensory holoprojections to derive associational information such as that concerning depth perception, for example (from both audio and visual sources in combination), as well as perception of motion, types and categories of perceptions, as well as cross-modal associations. Thus the primary holoprojections generate far more from the memory data than just elementary sensory information. This is why the ILN scan cannot be simply of the primary holoprojections themselves, but of the result of the entire sequential associative process they generate in the cortex. The process extends over the entire cortex. For instance, at the most advanced level of associational processes in the frontal cortex regions, you have a bound, unitary, and multi-sensory short-term-memory of events (and your reaction to these events) that just happened a moment ago, produced by successive stages of associational cortex operation. This memory information is also scanned and becomes part of the habit routine complex. In this sense even current experience is very much like a reverberation, for current evaluation of reality is based upon the interpretation of reality just experienced which has been re-injected into the ongoing habit routine complex. Thus the significances that thinking2 decides to examine more closely by extracting information from the primary holoprojections are actively perpetuated. Using this mechanism we can increase the proportion of "raw reality" in the current habit routine to override the "interpretation" of reality that would be supplied by the unmolested habit routine alone. And here is where the psychedelic experience comes in. The process which is so startlingly activated by psychedelic drugs, is the very same process that we can accomplish, if on a more limited level, as just described. Let us look closer at this overall process, for it involves the generation of additional composite holoprojections which have more to do with conscious thinking2 operations, including free will, than with automatic sub- or pre-conscious thinking1 processes and their holoprojections.
The nuclei of the thalamus are also important for the generation of these composite holoprojections, but at this stage, other brain nuclei become partners with the thalamus and cortex. Again, these older, more "primitive" parts of the brain act as drivers in dialog with the cortex to produce the composite informational fields. (19) Among the brain components taking part are the locus coeruleus and raphe nuclei of the brain stem, the amygdala, hippocampus, the basal ganglia and cerebellum, among others. It is certainly an ambitious statement to say that I will attempt to explain the role of some these brain areas, but here goes:
First let us consider the role of the locus coeruleus and the raphe nuclei, for it is the nerve pathways connecting these two brain areas with the cortex and with each other that are the primary site of biochemical action of the psychedelic drugs. It is with these nuclei that I must show how significance detection and the suspension of habit routine is accomplished. I would propose that the locus coeruleus is the master functioning body, the driver which through dialog with essentially all brain areas but particularly the thalamus, the raphe nuclei and the cortex, produces the composite holoprojection containing information about significance or salience not only in the environment, but also even in the ongoing processes of thought leading to ideas, opinions, etc. I will refrain from presenting a diagram of the connecting nervous pathways between the various brain areas cited as it is extremely difficult to do so meaningfully at this stage. Certainly, as I have already said, there are plenty of connecting pathways between all of these brain parts, there are rather too many, it would seem.
Now the statement that the connections between the raphe nuclei, the locus coeruleus, and their connections to the thalamus and cortex are the primary site of biochemical action of the psychedelic drugs is based quite solidly on recent brain research. (20) We know that LSD and other psychedelics, for example, exert powerful influence on the operation of neurons emitting and receiving at their synapses the neurotransmitter, serotonin. There remains considerable mystery as to how the drugs react with these neurons, whether they activate or inhibit serotonin receptors, which types of serotonin receptors are affected, and so forth. There is also considerable mystery as to how these affected neurons might bring about the overall psychological result. The first question remains, at the present state of research, very difficult to answer. But it is with the second question that I believe we should start, for there seems to be enough information now available to formulate a model.
It is generally agreed by neuroscientists today that the locus coeruleus acts as a kind of novelty or significance detector, its activation (in animals) has been shown to increase in response to stressful or noxious stimuli, preferred food and other complex arousing events, and even to changes in body systems such as the level of oxygen or carbon dioxide in the blood. The psychedelic drugs have been repeatedly shown to greatly increase the activity of the locus coeruleus, but not when applied directly to the cells which make up the nucleus. Thus it has been hypothesized that the state of the locus coeruleus must be influenced by another nucleus or system which itself is directly affected by psychedelics. Some relevant facts concerning the locus coeruleus:
The locus coeruleus (LC) consists of two dense clusters of noradrenergic neurons located bilaterally in the upper pons at the lateral border of the 4th ventricle. The LC, which projects diffusely to virtually all regions of the neuraxis, receives an extraordinary convergence of somatic, visceral and other sensory inputs from all regions of the body and has been likened to a novelty detector. Thus, the LC represents a unique nodal point both for the detection of significant changes in the internal and external environment and for relaying this information to the remainder of the central nervous system. It is not surprising that hallucinogenic drugs, which produce such dramatic changes in perception, would alter either directly or indirectly the function of LC neurons. (21)
In keeping with the holonomic model, I would propose that the locus coeruleus is the driver which produces a composite holoprojection consisting of the superimposition and canceling of primary holoprojections including the habit routine holoprojection to yield a field of information concerning significances in the ongoing experience of the organism. The detection of significance, or salience, normally is derived from the same memory data from which the habit routine complex is generated, and is merely a repeat detection of salience that has occurred in the past. When the test animal mentioned above is shown a preferred food, for instance, and its locus coeruleus is shown to increase in activity, the salience detected is obviously relative to memory data of the preferred food. But when I am the test animal in a French restaurant, when some unknown yet succulent dish is placed before me, I begin to extract information directly from the primary sensory holoprojections to try to deduce the composition and possible methods of preparation of the mysterious delicacy. Memory information in the habit routine will still be the primary source of information in these deliberations, but my Attention will guide the process to actual examination of the "genuine" sensory data contained in the primary holoprojections. As I mentioned above in introducing the functions of the primary holoprojections, we see that salience detection may use the primary data, but normally, and routinely, salience is merely a repeat performance of previous detection, based on the habit routine complex itself.
The connections of the locus coeruleus which accomplish the detection of normal salience from the habit routine data may be simply the interconnections with the cortex. The locus coeruleus receives a modest input from only one area of the cortex, the prefrontal cortex, but it sends its output to the entire cortex. We see again (as in the case of the pathways between the thalamus and the primary sensory cortex) the situation where the "return" signal is far more important than the "input" signal, and this suggests, as in the nervous pathways taking part in the generation of the primary sensory holoprojections, a reverberation, in this case the salience detection holoprojection. (22) The signal from the prefrontal cortex is the final stage of the entire process of association, so it will obviously represent the complex associational memory data of past salience detection that was experienced. The return pathways to all cortex areas might also be thought of as facilitating the cancellation or ignoring of all features of the habit routine except for the salient entities, so that the resulting holoprojection contains only information about these entities. Thus the Attention is directed to the salience which practically jumps out of its surroundings. The locus coeruleus has several inputs besides that from the prefrontal cortex, but these inputs function to modulate the holoprojection generation in various ways. A particularly important control of the process is accomplished by the raphe nuclei.
The raphe nuclei of the brain stem are particularly important to salience detection and to the psychedelic experience because they contain the great majority of neurons of the brain which use serotonin as a neurotransmitter. Some of the earliest work on the effect of LSD in the brain found that "LSD and other indoleamine hallucinogens...have potent, direct inhibitory effects upon serotonergic neurons located in the raphe nuclei of the brainstem." (23) Now it has been well established that the serotonergic neurons of the raphe nuclei project heavily to the locus coeruleus, and likewise that serotonin inhibits the firing of the type of neuron found in the locus coeruleus. (24) As mentioned above, studies have also confirmed the psychedelic agents have as their target at least some of the many types of serotonin receptors on neurons both of the areas signaled by the raphe nuclei, and on the raphe serotonin neurons themselves (autoreceptors). The raphe neurons also project widely throughout the brain, to all areas of the cortex, (strongly to the prefrontal cortex from which the locus coeruleus derives its input), the thalamus, the amygdala, virtually the entire nervous system. Thus I would propose that the raphe nuclei are the principle mechanism of control, the driver which guides the mode of function of salience detection, as well as the manner in which the detection is used and subsequently stored in memory. It appears that Attention and other thinking2 process use the serotonergic system of the brain, based in the raphe nuclei, to direct the detection and use of salience, but not simply by "inhibition" as would be implied by the observed "inhibitory" action of serotonin on the neurons of the locus coeruleus, or the observed "inhibition" of serotonergic neurons by LSD.
It is a curious fact of the human brain, that fully 75% of the neurons therein are supposedly "inhibitory neurons", whereas in the monkey the figure is 45%, and in the cat a mere 35%. (25) Clearly these facts must have some tremendously important significance, not only for the type of functioning they imply of the brain, but in some sense they must tell us something very important about the psychological differences between man and animals, particularly the great disparity of intellectual capacity. A not very convincing speculation has been that the large proportion of inhibitory neurons allows a "streamlining of thinking" in which groups of brain cells are more quickly returned to a state of readiness after some operation. (26) I would propose that the inhibitory neurons do not "inhibit" in such a literal manner, but rather are concerned with the superimposition of holoprojections in which a cancellation or subtraction of information results, such as the manner in which all peripheral information from memory going into the generation of the salience holoprojection is removed so as to yield a composite holoprojection consisting of only the detected entities. It might be said that human mental powers, as opposed to that of lower animals, reside primarily in the ability to discriminate between and detect widely differing types of significance not only in the environment but within thought patterns of abstractions and concerning ideas and constructs of the intellect. These human feats, I would propose, are accomplished using the wide network of inhibitory neurons functioning to produce composite holoprojections derived through the comparison and subtraction of informational fields one from another, to reveal patterns the complexity of which far outstrips the power of lower animals to detect.
There are probably several brain operations, brought about by combinations of brain parts including a nucleus of the midbrain or brainstem as a driver, which generate such composite holoprojections. The detection of emotional content, or valence, is probably accomplished using the amygdala as a driver in dialog with the cortex and other nuclei, superimposing the same primary holoprojections as are used for salience detection. But the salience detection of the locus coeruleus, controlled and modulated by the raphe nuclei connections throughout the brain, generates what is probably the most important holoprojection of the brain/mind, (or in any case, second only in importance to the habit routine holoprojection). And it is upon the generation of this salience detection holoprojection that the psychedelic drugs exert their effects.
Now it was soon noticed, after the inhibitory effect of LSD and similar psychedelic drugs on the raphe nuclei was discovered, that other psychedelic agents such as mescaline and the phenethylamine family did not produce the inhibition. The indirect effect on the locus coeruleus was, however, as important as with LSD. The effect is indirect, for as mentioned, the application of psychedelic agents to locus coeruleus neurons themselves fails to alter their activity. But since the locus coeruleus salience detection system involves the entire cortex, and the serotonergic neurons of the raphe nuclei project to the entire nervous system, it is evident that the control of salience detection would be alterable at many different sites of potential psychedelic drug action. A direct change in the raphe nuclei is the possible primary action in the case of LSD, whereas a change in the effect of the signaling by the raphe neurons, either in the locus coeruleus or possibly the cortex itself, might be the mechanism for mescaline. The overall effect is in both cases a change, which appears to be an increase, in the rate and type of salience detected by the locus coeruleus system. I am tempted to repeat some of Huxley's observations about significance quoted at the end of chapter 3, but instead will quote Alan Watts, here writing about his first experiment with psychedelic drugs:
"I have said that my general impression of the first experiment was that the "mechanism" by which we screen our sense-data and select only some of them as significant had been partially suspended. Consequently, I felt that the particular feeling which we associate with "the meaningful" was projected indiscriminately upon everything, and then rationalized in ways that might strike an independent observer as ridiculous—unless, perhaps, the subject were unusually clever at rationalizing. However, the philosopher cannot pass up the point that our selection of some sense-data as significant and others as insignificant is always with relation to particular purposes—survival, the quest for certain pleasures, finding one's way to some destination, or whatever it may be." (27)
I have suggested above that there are two modes of salience detection, the first merely automatic and based upon previously experienced situations, and a second which is based on the genuine data of the primary holoprojections. Remember my little story of noticing the grey rock in the middle of the pathway. Automatic salience detection brought it to my awareness, and interpreted it relative to previous experience. Yet some unconsciously perceived anomalous data, a slight movement, a color not quite in keeping with experience, caused me to suspect an error, and choose to examine the raw sensory data itself and Decide that something was amiss. At this point the original habit routine was overruled, and a new interpretation actively demanded by the Attention. This was accomplished by use of the working memory, thought to be a function of and resident in the prefrontal cortex, the same cortical area from which the locus coeruleus receives its sole input. Remember also my stated feeling that under the influence of psychedelics, it seems that the habit routines of interpretation in this case would be at least momentarily suspended, and after a moment multiple habit routines might arrive at thinking2 processes. All these observations seem to indicate that psychedelics would be interfering with the habit routine holoprojection, rather than the salience holoprojection. But according to my neurological model so far, it would appear that psychedelic agents interfere with the control exerted on the salience holoprojection by the raphe nuclei and the serotonergic neurons extending widely to all areas of the brain.
I would now propose that the profound alteration of the salience holoprojection by psychedelics, illustrated both by neurological data and by the practical observations of Huxley, Watts, and many others, causes the individual, using the conscious mechanisms of thinking2, to himself suspend the dependence on the habit routine holoprojection. It is the cumulative effect of not only added salience detected in the ENV both external and internal, but of the interpretation of this increased salience as itself extremely significant, that leads to a veritable avalanche of salience detection which simply overwhelms normal acceptance of and dependence on the habit routine system.
The habit routines are still assembled and are there in the background, but they are almost completely ignored by Attention, Decision, and other thinking2 processes amidst the flood of salience perceived. Thus, the psychedelic experience is, in an important sense, voluntary, and this would explain the ability to achieve such states through meditation and other voluntary mechanisms. That the habit routines are still assembled and available to some extent is illustrated by the common ability during the lucid end-stages of psychedelic experience to recognize a duality in experience: a perception of the way things appear with "the doors of perception" cleansed, and a simultaneous recognition of how the same scene would appear in a normal state of mind governed by habit routine. Such realization extends to the perception one's personality traits, one's prejudices and automatisms of behavior, from a viewpoint that is essentially outside of the self and beyond the ego. In this state I believe, the habit routines have been completely suspended in function, and salience detection is using the information of the primary holoprojections.
At the request of thinking2, the primary sensory holoprojections become the subject of meticulous examination, genuine reality floods through, the habit routines ignored. The raphe nuclei must in some sense be acting as a control mechanism for this switch-over, allowing the locus coeruleus system to create superimpositions of the genuine data rather than the memory data. This switch-over probably occurs in the cortex itself, by the control exerted by the serotonergic neurons from the raphe nuclei which contact both inhibitory interneurons and the pyramidal neurons of the cortex. (28)
Experimental evidence supporting the above model relating the neurological operations of brain systems and the habit routine search and suspension hypothesis of normal and psychedelic functioning has been easy to find in the literature. Of course, due to the current state of neuroscience, it is often found that alternative interpretations of experimental results, leading to radically different models, is possible. Such is the case here, and for any specific experiment which I might use as evidence for my model, others would find it just as easy to use the same data for another view. For this reason, as well as in the interest of brevity, I shall mention only a few examples. A thorough survey would require at least another volume, and the more important chapters of the present volume still await exposition.
A vast body of literature concerning brain function and its perturbation by brain lesions goes back more than a century. A great many studies have been done concerning patients whose brains have sustained damage through accidents or necessary brain surgery, and experimentally, countless numbers of animals of every description have undergone destruction, disconnection or removal of various brain areas in the attempt to localize various sensory, motor, and cognitive functions of the brain. With such a wealth of evidence, it is not hard to find studies that might support almost any model one would care to dream up.
I will mention just a few cases which have been well documented, and which deal with damage to the areas of the prefrontal cortex. This brain area, it will be remembered, has been suggested both in the literature and in my own model as an important center involved with the working memory and with the most complex levels of associative processing by the cortex. Such association might be expected to facilitate complex cognitive phenomena such as the expression of personality traits, decision making and attention, voluntary action and free will, the perception of and reaction to complex social situations, i.e., the most complex and human of cognitive functions. (It is the frontal lobe, and especially the prefrontal area that has seen such a massive expansion and development in the recent evolution of the hominids.) Remember also that it is from the prefrontal cortex that the sole cortical projection to the locus coeruleus occurs. Projections from the prefrontal cortex also extend to the raphe nuclei and the amygdala.
The model I have devised would therefore predict that disruption of frontal lobe integrity should produce alteration of salience detection and the evaluation and expression of emotion, and radical changes in the assembly and use of habit routines, especially as they apply to these most advanced cognitive and affective functions. Since the connections to the locus coeruleus, raphe nuclei, and amygdala hypothesized to be important in my model project from the prefrontal cortex, interference with salience and valence functions should certainly be observed in cases of prefrontal damage. And prefrontal damage resulting in an impaired function of working memory in supplying parameters for the ongoing habit routine search process should produce symptoms identifiable as resulting from impaired generation, access to, or use of habit routine in ongoing cognitive operations.
A very famous case of brain injury in the prefrontal area, recently the subject of a book and various newspaper articles, is that of the construction foreman Phineas P. Gage. In 1848 Gage sustained a massive brain injury when an explosives procedure went terribly wrong and sent an iron tamping bar vertically through the frontal region of his brain. Miraculously, Gage seemed at first practically unaffected, even walking some distance, and conversing with his men on the way to medical attention. It was only later that the peculiar kind of mental deficits that necessarily result from this type of prefrontal injury came to light. The case has recently become the subject of detailed attention thanks to the work of Hanna Damasio, who was able to reconstruct the precise location of Gage's brain damage using state-of-the-art computer techniques to analyze the features of Gage's skull, a museum exhibit at the Harvard Medical School Museum for over a hundred years. An overview of this work together with a detailed examination of the symptomatic evidence in the case is presented in a recent book. (29)
It was said of Gage that he became a different man, his entire personality was radically changed. It seemed that he had completely lost access to his previously acquired social conventions and rules and had become childish, irresponsible, yet strangely undiminished in terms of episodic and autobiographical memory, language ability, even, one might say, intelligence. In terms of the habit routine model: In the daily yet complex social relationships in which personality is expressed, it appeared that Gage's behavior was capricious, or even random, the habit routines of personality which are among the most complex and highly associative aspects of the habit routine complex, were no longer accessible (reconstructible) from the memory of the frontal cortex and by consequence, his automatic social behavior was based not upon previous experience but was instead arbitrary and random rather than merely childish. In theory, it seems, he could still reason out how one should act in a given situation, but when forced into a decision by a life situation, he was essentially powerless to apply such reasoning. This may illustrate how dependent we are on habit routine for making decisions and for the expression of personality. In the case of personality traits, it would obviously be impossible to calculate logically and consciously how to react to a situation in accord with our established personality. The reaction must be automatic and instantaneous. If this is also the case with at least some aspects of decision making, a statement I previously made becomes even more pertinent: "We see what we have already seen..." and decide in ways that we have previously decided to an overwhelming degree.
The symptoms described by Antonio Damasio (30) could well be explained in these terms, but in addition, Damasio describes the case of a prefrontal patient which he had himself examined in detail. The patient, referred to as Elliot, had undergone surgery for the removal of a tumor at the base of the frontal cortex, just above the eye sockets. Surrounding brain tissue had also been removed or damaged, and thus a large portion of the prefrontal cortices were dysfunctional. As with Gage, the largest part of the damage was in the ventromedial (lower-central) area, and many of the symptoms were repeated, such as radical personality change. But particularly striking were symptoms that might be interpreted as resulting from interference with the salience and valence systems involving connections from the damaged prefrontal region to the locus coeruleus, raphe nuclei, and to the amygdala. These symptoms might be categorized as a lack of ability to plan ahead, to make decisions concerning strategy and the immediate future, almost, one might say, a deficit of free will. The obvious intelligence which Elliot retained could not be mustered to organize even simple sequences of activity as required for his job, for instance. In addition, he exhibited an emotional flatness or detachment in striking contrast to his pre-operative character.
Interestingly, a lengthy series of psychological tests indicated that, like Gage, Elliot could theoretically make such decisions, such as those pertaining to moral judgments, ends and means problem solving, awareness of consequences concerning both events and social situations, etc. But when a real life situation forced a decision based on habit routine, the theoretical knowledge seemed impossible to apply. In the theoretical situation, we could say that the records of social information in memory were actively and intentionally used to reconstruct the required decision from scratch, whereas in the life situation what was required was an automatic referral to the habit routines representing such decisions that had been made over a lifetime: the records of previous decisions as represented in the highest levels of habit routine had been destroyed along with frontal cortex, (31) whereas the previous memories themselves upon which the previous decisions had been based were still intact in other areas of the cortex.
The combination of decision deficit with emotional flatness led Damasio to construct a model he calls the Somatic-Marker Hypothesis, and it has attracted much favorable comment. (32) I will not describe it here, but will instead offer my own interpretation of the concurrence of the two symptoms. We could describe an inability to make rational decisions not only as due to deficits in the assembly of the highest levels of the habit routine complex, but also on the basis of faulty salience detection, since the relative significance of events and aspects of reality, both present and in memory, must obviously play an important role in constructing plans based upon contingencies and the evaluation of probabilities and strategies. With both of the above cases, Gage and Elliot, there was a theoretical ability to make decisions, based upon reasoning processes utilizing episodic and autobiographical memory of events themselves. Likewise, reasoning processes and memory would be able to deduce significance both in the internal and external environments, but the automation of salience detection would be deficient. The key to understanding the overall syndrome, however, is seeing that the generation of emotional content by a holoprojection driven by the amygdala must be a process based on information in the salience detection holoprojection. It is with the various significances detected in the external and internal environment that emotional expression deals, all routine and superfluous information merely falls by the wayside and is ignored. The SD holoprojection feedback to all areas of the cortex accomplishes this cancellation or ignorance of irrelevancy, to leave the detected significances in stark contrast relative to the background. From this holoprojection of detected significance comes the data from which the emotional value or valence is generated.
It is easy to see, then, that if the salience detection system is perturbed or interrupted, a naturally resulting symptom should be emotional flatness, or even randomly expressed emotion since the amygdala is not itself damaged but merely has little or no accurate information to work with. Elliot himself realized perfectly well not only his inabilities in making decisions in the face of real-life situations, but also how subjects or situations that had once caused him strong emotion no longer evoked any reaction whatsoever. Here we can see that, since he remembers subjects which formerly caused an emotional reaction, he can also theoretically evaluate emotional content just as he can theoretically evaluate significance, reconstructing the information from long-term memory. But it is the automated accomplishment of these functions which has been perturbed, and in real-life, on-the-spot decision making, reconstruction does not and cannot substitute for the automated processes.
According to Damasio's hypothesis, the deficit for decision is based on the deficit of emotional content, but I believe the situation is quite the reverse, that emotion is based on significance detection necessary in the process of automatic decision-making, and that both of these functions are based upon intact operation and connections of the prefrontal cortices to the locus coeruleus and the raphe nuclei.
Now I cannot tell from Damasio's descriptions whether the actual connections projecting from the frontal cortex to the locus coeruleus, amygdala and raphe nuclei were damaged or severed during Elliot's operation. It would seem in Gage's case that the trajectory of the iron bar might well indicate that these connections were destroyed: they are grouped together into the medial forebrain bundle, an important nerve pathway passing directly through the ventromedial area. This pathway also contains the projections returning from the locus coeruleus and raphe nuclei which connect to all areas of the cortex. Whether the severing of the medial forebrain bundle alone produces a syndrome similar to the actual destruction of prefrontal cortex is not known, but if my hypothesis is correct, the severing of the medial forebrain bundle connections to the locus coeruleus and raphe nuclei should produce a very similar result as is seen in cases such as Gage and Elliot. Damage to the frontal cortices may have to be quite widespread to bring about the same result as the simple severing of these nervous pathways.
There is another type of damage to the frontal cortex which produces rather different results. So far, in the two cases mentioned, the principal damage was to the ventromedial areas, just above the eyes and centrally located. When damage to the dorsolateral areas also occurs, psychological tests indicate an important deficit in working memory accompanies the syndrome. These same tests, given to Elliot, showed no disability whatsoever in his working memory function. Inasmuch as the working memory has been proposed here as an important part of the process of habit routine search, its disruption should alter the process in certain ways. In the cases of ventromedial damage cited above, I proposed that the actual memory information necessary for construction of the highest associative levels of the habit routine complex had been destroyed. But with dorsolateral damage as well, an important part of the system which carries out the habit routine search is destroyed: the ability to supply parameters for the search is impeded.
It was proposed that conscious and unconscious parameters guiding the successive scans producing the habit routine complex were introduced via the working memory. The ILN scan, incorporating WM parameters would, on the succeeding scan retrieve a modified selection of memory information according to the parameters previously scanned. And it was proposed that this "small window on reality" was essentially the only normal way to guide the processes of thinking1 using free will or intentional creativity. In experimental situations we notice deficits in precisely these domains. Fuster, in his book on the prefrontal cortex, states that "In general terms, ablation studies indicate that the cortex of the dorsal and lateral prefrontal surface is primarily involved in cognitive aspects of behavior. The rest of the prefrontal cortex, medial and ventral, appears to be mostly involved in affective and motivational functions..." (33) (such as salience detection and emotional evaluation.)
In human prefrontal patients, a striking experimental demonstration of working memory disruption due to dorsolateral damage is the Wisconsin Card Sorting Test, here described by Dudai:
The subject is presented with a series of stimulus cards and a deck of response cards. The cards bear coloured geometric patterns (e.g. a single blue star, three red circles), and can be matched by categories (e.g. colour, form, number). The examiner selects a sorting category (e.g. colour), but does not inform the subject. The latter is instructed to place a response card in front of a stimulus card, wherever he or she thinks it should go. The examiner then informs the subject if the response was right or wrong, and the subject uses this information to obtain correct responses in the following matches. After ten consecutive responses, the examiner shifts the sorting category without warning, and the subject must unveil it again to obtain correct matches. The procedure is then repeated with other sorting categories. Patients with prefrontal lesions find this task abnormally difficult. The interpretation is that they have difficulties in using temporarily stored information to regulate their actions. (34)
The function of working memory as a parameter store for ongoing habit routine search is well illustrated by the experiment. The original instructions for the experiment, which the subject has little difficulty in following, create a simple habit routine for performing the sorting according to the first learned category. But when the category is changed, the habit routine developed for the experiment remains fixed, its alteration by new working memory parameters is difficult if not impossible. Most prefrontal patients with dorsolateral damage have great difficulty in this test, but Elliot, whose damage was limited to ventromedial areas, passed it with flying colors.
Now the evidence concerning prefrontal damage and its interpretation using my model takes on some additional relevance in consideration of the following experimental findings: In a series of experiments using PET brain scan techniques to observe subjects in altered states of consciousness brought about by psychedelics, the primary effect noted was a significant increase in the activity of the frontal cortices. (35) This result, in combination with Aghajanian's findings of greatly increased locus coeruleus activity caused by psychedelic drugs, lends my interpretation some credibility. Under the influence of psychedelic drugs, the cognitive functions of working memory and habit routine search, significance detection and emotional value detection are all working overtime, and they all are facilitated by the prefrontal cortices. Whether this is all mere coincidence, or an indication that pieces of a very intricate puzzle are falling into place only time and further research will tell.
A vast quantity of experimental evidence awaits the organizing ability of some yet-to-be-discovered overall model of brain function. In reading the many papers dealing with just the prefrontal cortex in the recently published The Cognitive Neurosciences (36) for example, one is immediately impressed with both the wealth of experimental information available and the corresponding wealth of models, terminologies, and hypotheses which attempt to organize this information. But such a cornucopia of viewpoints must certainly be a sign that we modelers are very much like the collection of blind men describing the elephant from the feel of merely local areas of the overall beast. Who will be the visionary to discover the viewpoint from which all these models and observations become a united whole? I certainly cannot pretend that the cognitive and neurological models I have presented here fulfill that function. I would be the first to admit the highly speculative nature of the above neurological model that I have presented, and would not be surprised nor indignant if it were said that the area that I was describing was not even part of the elephant!
In my view, the only claim for consideration of my ideas stems from their origin in the attempt to explain the body of evidence that has accumulated concerning the psychedelic experience, evidence which has been almost entirely disregarded by the mainstream of science for nearly thirty years. Had it not been for such neglect, which in part was forced by an idiotic international effort to fight an unwinnable, self-defeating and therefore irrational "war on drugs", it seems to me that several fields of study of human psychology and neuroscience would have by now achieved far greater insight than is the case.
But we can blame not only the drug warriors, the politicians and intelligence organizations, the religious moralizers and puritanical oafs for this ignorance: scientists too are to blame, perhaps equally so. It was easy for me, from the outside of the scientific establishment, to see that one of the most important discoveries ever made by Western science was being ignored, even vilified. But from the inside of that scientific enterprise, it was apparently no easier to see what was happening than it was for those inside the traps of religious fanaticism or the carefully cultivated paranoia which is the paradigm for institutions providing much of the raw material for the politicians: the intelligence organizations.
Scientists, at least outside the realm of their own specialties, often seem as prone to narrow-mindedness as are other intelligent yet confused men. It seems to be almost instinctual that men follow such narrow pathways through life, and the habit-routine model is certainly also an attempt to show how such narrowness might actually be derived from an inherent neurological feature of the human organism, rather than something which we must label instinctual for want of a better understanding. The functioning of our nervous systems utilizing the habit routine system might be taken as a convenient excuse for the current deplorable state of civilization, this "century of holocaust," but now that I have laid bare the roots of the situation, it is a lame excuse at best. In the next chapter I will explore the idea that this inherent neurological and cognitive feature has been an essential (yet now for man a skeuomorphic or vestigial) characteristic in the stages of evolution not only of man, but of all animal life.
References
(1) Rupert Sheldrake, The Presence of the Past, 1988, William Collins, publisher. (back)
(2) A brief but concise description of the Hopfield Network may be found in Crick, The Astonishing Hypothesis, ibid., pp182-185. A more technical and thorough exposition is found in Churchland & Sejnowski, The Computational Brain 1993 MIT Press, p82ff. (back)
(3) Steven Rose, The Making of Memory, Bantam Press (Great Britain) 1992, see chapter 13. (back)
(4) Ibid., p322. (back)
(5) Karl H. Pribram, Languages of the Brain, 1971 Brandon House, New York, recently reissued (1988). See also his essay "What the Fuss is All About", in The Holographic Paradigm, Ken Wilber (ed.), New Science Library, Shambala 1982. Other articles in this volume discuss the possibilities and limitations of the model. (back)
(6) "From Metaphors to Models: the Use of Analogy in Neuropsychology", Karl H. Pribram in Metaphors in the History of Psychology, David e. Leary, ed., 1990, Cambridge University Press. (back)
(7) Sometimes called the "grandmother cell" model, in which a unit of learning or recognition (of your grandmother, for instance) was supposedly associated with a single brain cell or strictly local set of interconnections between cells. (back)
(8) Francis Crick (1994) ibid. p185. (back)
(9) Introduced by George Leonard to refer to entities having the nature of a hologram, The Silent Pulse, Dutton, New York 1978. (back)
(10) "Spectral Density Maps of Receptive Fields in the Rat's Somatosensory Cortex" in Origins: Brain & Self Organization, Karl Pribram, ed., 1994, Lawrence Erlbaum Associates. (back)
(11) Brain and Perception: Holonomy and Structure in Figural Processing, Karl H. Pribram, 1991, Lawrence Erlbaum Associates. (back)
(12) Times Literary Supplement, London, August 25, 1995. (back)
(13) See "Quantum Coherence in Microtubules" by Stuart R. Hameroff, Journal of Consciousness Studies 1, No.1, 1994 pp91-118. (back)
(14) In the brain there are approximately 1011 neurons (100,000,000,000), 1015 synapses, but Hameroff estimates that there may be 1023 dynamic sites or states associated with the microtubules. (back)
(15) See "Perception as an Oneiric-like State Modulated by the Senses", Llin�s and Ribary, in Large-Scale Neuronal Theories of the Brain, 1994, MIT Press. On page 113 is a summary of brain connectivity illustrative of the inadequacy of current "information-flow" models. (back)
(16) In producing a holographic image by illuminating a holographic photographic plate (the hologram) with coherent light, the same image is produced by directing the light through only a small part of the plate as is produced by illuminating the entire plate. But the image in the former case carries much less definition, it is of lower resolution. (back)
(17) A review of the experiments is in Human Memory, Theory and Practice, Alan Baddeley pp14-18. (back)
(18) See "Perception as an Oneiric-like State Modulated by the Senses", Llin�s and Ribary, in Large-Scale Neuronal Theories of the Brain, 1994, MIT Press. (back)
(19) I might go so far as to suggest that most, if not all cognitive functions of the brain are accomplished by a dialog among two or more brain parts, and not by a single area acting alone to effect some cognitive result which is then passed on to another area. The multiple holoprojection model is entirely in accord with this suggestion. The model has conceptual similarities to another important "checks and balances" feedback system of the body, the hormonal system. (back)
(20) A publication containing the most recent research findings of the important workers in this field is 50 Years of LSD: Current Status and Perspectives of Hallucinogens, Pletscher and Ladewig, editors, Parthenon Publishing, 1994. The book presents papers submitted to a symposium of the Swiss Academy of Medical Sciences in October 1993. (back)
(21) Ibid., "LSD and phenethylamine hallucinogens: common sites of neuronal action", G.K. Aghajanian. (back)
(22) Another way of thinking about the disparity between "input" and "output" signals again illustrates a basic conceptual fault with the computer model of brain function. In the case of the locus coeruleus for example, it has been stated that since its "output" extends to such diverse regions, its functions must also be multiple and widespread. This view has the underlying assumption that the locus coeruleus is sending information it has processed from its modest input, to many locations where this data is then used for many different functions. But the alternative view I have proposed is that the locus coeruleus accomplishes only one function. The multiple and widely connected "output" pathways are not to be seen as sending information, but rather as requesting or accessing information of a widely diverse nature (relating to the detection of salience in many domains, situations, and complexities). The result of this request is then cycled back to the locus coeruleus via its modest input from the frontal cortex, as a reverberating holoprojection. (back)
(23) Ibid, p27. (back)
(24) Chemoarchitecture of the Brain, Rudolf Nieuwenhuys, 1985 Springer-Verlag, p40. (back)
(25) From references mentioned in Stairway to the Mind, Alwyn Scott, Springer-Verlag 1995, p94. (back)
(26) A suggestion of Donald Hebb, recounted in Stairway to the Mind, ibid., p94. (back)
(27) "The New Alchemy" in This Is It, Alan Watts, Random House, 1958. (back)
(28) Aghajanian in 50 Years of LSD, ibid., pp33-34 (back)
(29) Descartes' Error, Antonio R. Damasio, G.P. Putnam's Sons, 1994, chapters 1 & 2. (back)
(30) Ibid. (back)
(31) One could view the situation also as an inability of the intralaminar nucleus of the thalamus to include in its habit routine-generating scan the information from the destroyed frontal region. This is probably the more useful if not accurate view, as opposed to the view that frontal lobe "information" has been destroyed. (back)
(32) Ibid., p173-ff. (back)
(33) The Prefrontal Cortex, Joaquin M. Fuster, 2nd edition 1989, New York: Raven Press, p74. (back)
(34) The Neurobiology of Memory, Yadin Dudai, Oxford University Press 1989, p263. (back)
(35) F. X. Vollenweider and colleagues in recent papers summarized in "Evidence for a cortical-subcortical imbalance of sensory information processing during altered states of consciousness using positron emission tomography and [18F]fluorodeoxyglucose" in 50 Years of LSD, ibid., pp67-86. (back)
(36) Michael S. Gazzaniga, editor, The Cognitive
Neurosciences, 1995 The MIT Press. (back)
(note) Update to the 1996 edition of The Center of the Universe: The proposal that background neural activity is not mere random noise is supported by recent findings indicating that background neuron firing is fractal in nature. In the Journal of Neuroscience (vol 17, p 5666) Malvin Teich of Boston University notes that the average release rate [of neurotransmitter packets] fluctuates as dramatically from minute to minute as from second to second. Such repetition at different scales is a hallmark of fractals. (Quotation from New Scientist, 16 August, 1997.)(back)