">IMMUNOLOGY AND GLOBAL BEHAVIOUR
By
Professor Madeleine Bastide
Emeritus
Professor
Laboratory
of Immunology and Parasitology
Faculty
of Pharmacy, University of Montpellier I
2 - DYSFUNCTION
OF THE IMMUNE SYSTEM
3 - THE IMMUNE
SYSTEM IN THE INDIVIDUAL'S GLOBAL AWARENESS
4 -
COMMUNICATION LEVELS OF THE LIVING:
5 - NEW
INTERPRETATION OF THE PATHOLOGIES OF THE IMMUNE SYSTEM
AND THEIR
THERAPY
6 -
REFERENCES
The immune system always fascinates with its complexity and originality.
Indeed, its distribution throughout the organism by the lymphatic system and the
lymph nodes, its presence materialised by various organs with apparently
different functions (bone marrow, thymus, spleen, liver, etc…), and especially
its role, connected first of all to the phenomena of cell communication, make it
an ideal subject to illustrate the concept of global awareness. This system has
been called the “the mobile brain”, which implies a function way beyond the
classical mechanistic concept and suggests a role with an “informational”
capacity.
After quickly recalling the adaptation and the working of the immune
system, we are going to analyse its dysfunction resulting in various
pathologies, comparing the vision of this system within the classical
mechanistic concept with the one achieved in another thought structure. If we
depart from the vision of molecule-receptor interactions towards another model
integrating the immune system into the individual’s global awareness, into the
significant interpretation of its relation with the outside world, we notice
that pathologies (and their therapies) allow a new
interpretation.
Several unconventional therapies, far removed from the mechanistic
allopathic therapy, seem especially well adjusted to correcting the dysfunctions
of the immune system.
I –
FUNCTION OF THE IMMUNE SYSTEM
The distinction between “the self” and the “different from the self” is
the basic principle of the immune response. The difference from the self (commonly
designated
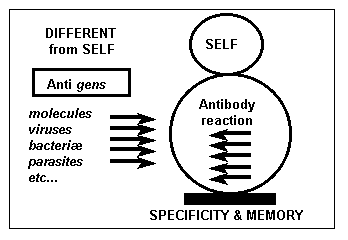
Figure 1:
Immune defence
by the term “non self”, which implies its
identification…) can be identified by the organism which triggers the
elimination systems of what is different: it is the antibody reaction
(figure 1).
The
self is represented by molecules of a glycoproteinic nature depicted on the
membrane of all the cells of the system and designated by “Major
Histocompatibility Complex” or MHC. Indeed, the diversity of these molecules
from an individual to another is such that they are able to characterise the
individual: they represent the self and take part in the rejection of a
transplant based upon the difference of MHC between the donor and the
receiver.
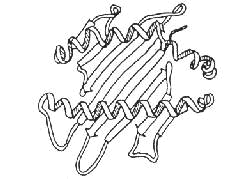
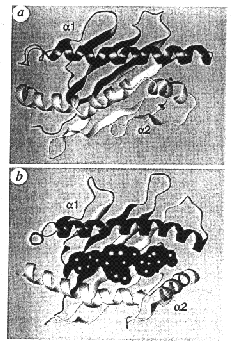
Figure
2: Structure of the MHC
molecule
(class I or II) presenting
a
pleated rigid part b
and two helices a
between
which the peptide is inserted
These
molecules belong to the super-family of immunoglobulins. The homologies
appearing between the structures of the molecules of this important family imply
that they derive from an ancestral structure (going back to about 500 million
years) which, throughout evolution, has remained a basic genetic and molecular
unit. In this molecular family we find recognition molecules of the immune
system (i.e. the antigen receptor of the T lymphocyte or TCR, the antigen
receptor of the B lymphocyte or immunoglobulin), at the top of the hierarchy of
adaptation to diversity, then MHC molecules which play the role of “marker” of
the self and “presenting” peptide molecule to the T lymphocyte receptor. Indeed,
we know now that the MHC has a peculiar structure accommodating a pocket inside
which peptides originating from the antigen extraneous to the system will settle
(figure 2). The union MHC + extraneous peptide is placed at the surface of the
antigen presenting cell, ready to be recognised by the T lymphocytes using the
antigen receptor TCR (Figure 3). It was demonstrated by crystallographic
analysis (Bjorkman, 1987) that this pocket is always filled : either by peptides
of the self unable to trigger a T lymphocyte response or by peptides different
from the self (antigen), able to trigger the T lymphocyte activation (Figure 3).
In this immunoglobulin family, we also find many molecules such as cell adhesion
molecules, immunoglobulin receptors, but also neurone adhesion molecules,
confirming the original relation between the nervous system and the immune
system in evolution.
The
triggering of the immune response is connected to this sole recognition of the
difference of the self. As soon as this recognition occurs, outpourings of
biochemical occurrences take place inside the T lymphocyte, resulting in its
activation by derepression of growth factor genes (interleukin 2 or 4). The
fixation of interleukin 2 on its receptor depicted by the T lymphocyte (for
instance T CD4 – TH1 lymphocyte), within an autocrine process, is going to
trigger the synthesis of other cytokines able to provoke the action of cytotoxic
T lymphocytes CD8 which are going to destroy the cells carrying the antigen
through a phenomenon of cytotoxicity (cell response). The TCD8, still designated
by the name of cytotoxic lymphocytes or CTL’s, will be helped in their
elimination of unwanted cells (target) by a cytokine, the tumor necrosis factor
(TNF), and by the joint action of two other cell types, the Natural Killer cells
and the "fully activated macrophages". These latter are going to produce
nitrogen monoxide (NO) which they release and, scattered over the target, kills
it. If the orientation of the response goes the way of the secretion of
antibodies (immunoglobulins specific of the antigen detected by the B lymphocyte
for a “shape” recognition), the activated TCD4-TH2’s release the cytokines
activating the B lymphocytes, which, under the action of various cytokines, are
going to evolve towards a plasmocyte-type structure and secrete profusely the
immunoglobulin with antibody function which had enabled their selection by the
antigen.
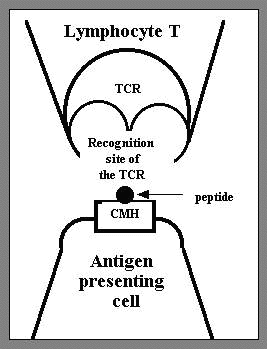
Figure
3: The recognition system
of
the peptide inside the MHC by the T lymphocyte.
The
receptor of the T lymphocyte antigen
recognises
the peptide and the MHC simultaneously
The special features of these two types of responses are the specificity (with regard to the
inductive antigen) and the set-up of a cell-type memory by maintaining the specific
clone.
The immune system ensures continuously the survival of organisms in an
environment where aggressors are multiple and diverse. Its regulation has a
special importance since the balance is worked out between an activity enabling
the defence with regard to extraneous elements while preserving the integrity of
the self. For this reason, there are regulations at all levels, producing a
subtle balance between the various cells, the cytokines (for instance interferon
gamma and interleukin 4 regulate each other), soluble receptors, regulating
antibodies (anti-idiotypic), hormones (the major negative regulator is
cortisol), neuropeptides and neuromediators (negative regulators of lymphocyte
activation such as vaso-intestinal peptide or VIP, calcitonin gene-related
peptide or CGRP, etc…).
II –
DYSFUNCTION OF THE IMMUNE SYSTEM
When, for various reasons, the balance of the immune system is disturbed
(stress, viral pathologies, immunosuppressor treatments, etc…) various
pathologies appear and show themselves in different ways according to the
subject’s genetic proneness, his environment, the nature of the immunological
fracture, etc…
Pathologies may be classified into three major groups, if we omit
congenital deficits originating from a genetic anomaly and the problem of grafts
and transplants. The latter represent a totally artificial situation of
dysfunction of the immune system created by a therapeutic act, the replacement
of an organ by another organ of another individual, of the same species or
not.
The first group includes pathologies due to an inadequacy of
efficiency of the immune system in the rejection of the extraneous aggressor
(possibly resulting in death). This phenomenon can be slight or affect the whole
system : the most suggestive case is the one of viral infections affecting
TCD4’s, like the action of HIV in AIDS. In this kind of pathologies, one tries
either to stimulate the immune system (immunomodulators), or to eliminate
pathogenic agents (anti-viral, antibiotics, anti-parasitic), or to eliminate the
cause (ex: anti-retrovirus in AIDS associated with a protection therapy against
the previously mentioned pathogenic agents).
The second group includes pathologies due to a “hypersensitivity” of
the immune system, the most widespread of which (30% of the population) is
immediate hypersensitivity or allergy. This hypersensitivity results
theoretically from an overproduction of immunoglobulins E which, fixing in
profusion on the blood’s basophil granulocytes or the tissue’s mastocytes are
going to provoke the degranulation of these cells every time that specific
antigens combine with the antibody site. The result of the degranulation is the
production of various inflammatory substances released from membrane
phospholipids, the formation of pro-inflammatory cytokines, of various
chemotactic substances, all this maintaining the pathology. There are two types
of therapies : either with the purpose of preventing or neutralising the
degranulation effects, or by injecting small doses of the allergen in question
to provoke a “desensitisation”.
It
is said that the allergic patient “sees danger when there is none” or that he is
“hypersensitive” to certain antigens.
The third group includes the auto-immune pathologies in which one
observes an immune response directed against certain molecules belonging to the
organism (auto-antigens) causing lesions self-maintained by the permanent
presence of the auto-antigen inside the organism. The true cause of auto-immune
diseases is unknown. However, one suspects viral or bacterial infections as
inductors of tolerance rupture to certain auto-antigens. They could be distorted
by the elimination of germs at tissue level by the immune response itself
(hepatitis and anti-liver response, juvenile diabetes and anti- pancreas
b
cell response, etc…). There are also “cross reactions” resulting from a
relationship of structure between the aggressor’s antigens and auto-antigens
(Streptococcus A and heart antigens, etc…). The “genetic proneness” connected to
the MHC structure, certain alleles of which enable a better presenting of
auto-antigens exist (the best example is the relation HLA B27 and the relative
risk of spondylarthritis, etc…). Then, there is the unknown, the exceptional
circumstances, “stress” and its relation to the immune system through the
hormonal ACTH-cortisol pathway or through the pineal gland and melatonin,
etc…
It seems to us that we can group together:
(i)
circumstantial auto-immune diseases (cross reactions, infections, etc…) the
appearance of which truly results from an error of recognition of the immune
system which is abused and which functions with a “mechanical”
error.
(ii)
“personality” auto-immune diseases, deeper, in which we can find certain
rheumatoid arthritis, multiple sclerosis, lupus erythematosus and all
connectivities, organ diseases such as thyroiditis (Hashimoto, Basedow) etc…
about which we know that they are often triggered after a violent stress, or an
immune unbalance, and the etiology of which is unknown. It seems to us that, in
this case, there is a grave error of interpretation between the self and what is
different and to be rejected, the process of central tolerance of the self being
disturbed : it is more a question of an “immunological” and/or physiological
and/or general loss of personality than a mechanical cause. This interpretation
corresponds to therapeutic approaches obtained with certain unconventional
therapies which treat the patient much more than the illness : indeed, the
allopathic approach is, in this case, restricted to palliatives,
immunosuppressors and anti-inflammatory medication, with the function of
“limiting the damage” without eliminating the cause.
III – THE
IMMUNE SYSTEM IN THE INDIVIDUAL’S GLOBAL AWARENESS
The term “global awareness” may not be translated by a sum of
interactions the way they are described in the classical notion of immune
response. Its definition calls for no anatomic reality and no localisation.
Global awareness is a dynamic property which is intrinsic to the living and may
not be reduced to its biological components. It is situated above an
interdependence of all systems leading to a modification or adaptation of the
body as a whole. The organism calls for any usable regulation process and mixes
both physical phenomena and psychic modification. Global awareness implies a
general management of the problems which are then going to be subject to
regulations adapted to each aggression. It translates the interactions within
the internal world and with the outside world, which are the characteristics of
the living. It is certain that the immune system is one of the tools enabling
the organism to withstand certain aggressions. It is certainly a very
accomplished physiological component which has been ensuring the perenniality of
species amongst hostile situations and aggressions of all sorts, the most
important one being microbial parasitism, i.e. the invasion by all viral,
unicellular or multi-cellular organisms developing to the detriment of a living
organism. The living is not a juxtaposition of mechanical systems based upon the
dogma of "all molecular”: the organism has the characteristics of the living,
its creative ability, its adaptability, its capacity of integration into the
environment and its temporality.
It would seem that the result obtained by the functioning of the immune
systems makes up an individual’s “global” function. But, for this, the
conceptual tools enabling this approach are lacking, for nothing has been
proposed to ensure the communication between soma and psyche, presenting
themselves, however, as an integral part of global
awareness.
I – Body signifiers as a mediation
between soma and psyche
The only possible mediation between soma and psyche is suggested by A.
Lagache (Lagache 1988, 1997a, 1997b). We owe her the suggestion of a new
paradigm, the one of body signifiers (figure 4) putting analogical communication
into play, enabling non-symbolic communications at body level. “The body and the mind are not objects,
neither are they closed “boxes” on their substance, but living information
networks, organised according to regular and intelligible
laws”.
The
information is going to use mimetic presentation to designate its object.
Everything happens in the form of a purely sensitive communication the support
of which may be implied. The only example of support we could give is the one
provided by homeopathic dilutions ; we know that they are sensitive to
electromagnetic radiations (Hadji & coll., 1992) or that their power is
transferred by adapted devices (Endler & coll., 1994, 1995, 1997). Why not
suggest as a hypothesis that this body information be transmitted by very low
intensity and very low frequency electromagnetic radiations emitted in a very
stable manner by succussed aqueous solvents. Water, which represents 90% of our
body, is succussed continuously by heart contraction at about 90 beats a minute.
It was recognised that, in a non-traditional manner, water plays the role of a
ligant of macromolecules, which confers it a mediation function of molecular
contacts (Douzou, 1994). This mediation function could also be situated at the
information level by endogenous electromagnetic emission which would thus carry
the information of the substances belonging to the organism. This could be the
case of certain cytokines such as the “Colony Stimulating Factors” (CSF’s), for
instance interleukin 3 released at very low concentrations in the
microenvironment of immunocompetent cells in the course of cell interactions
during the immune response. These CSF’s can have a long-distance effect on the
hematopoietic strain cells of the bone marrow (endocrine effect). This
phenomenon cannot be explained by a serous rate comparable to that of hormones,
because of the very low initial concentration after local release and the
dilution effect of the circulating blood. A hypothesis would be the
communication to the hematopoietic strain cells of the marrow by the “informant”
nature of these cytokines further to their blood succussness. The information
received by the body play the role of “biological or body signifiers”, able to
provoke physiological modifications after treatment of the information by the
organisms. This communication follows very specific rules different from
exchanges of objects (Bastide & Lagache, 1992, 1995,
1999).

Figure
4: the paradigm of body signifiers,
a
mediation between mechanistic and symbolic paradigms.
2 – The Immunological Self in the
paradigm of Body Signifiers
We can now introduce a new dimension in the concept of the Immunological
Self, which, let us recall, plays a fundamental role in the identification of
what the organism must reject (see § I). The Self and the global awareness of a
system are completely connected : indeed, global awareness rests upon the notion
of “self”, this “global” entity which defines a living structure, with a
complexity which is going to increase with its place within the scale of
evolution. Of course, we are concerned with the human system, the most perfect
in evolution, therefore the most complex, with its three levels representing the
paradigms indicated previously : the molecular level with its complexity and its
systems of cybernetic type retro-control ; the level of biological information,
with a carrier we can suggest as being of the electromagnetic type, and the
psychic level in its entirety. From the internal standpoint, the organism
communicates continuously, with exchanges at all levels, from the psyche to the
molecular (somatisation, allergies, etc...), building its body representations
in response to biological information (symptoms, etc...) (Bastide & Lagache,
1999).
The system also communicates permanently with the outside world. The
organism which receives an information or a stimulus on a given level and which
can respond to it on the same level adapts without any risk of pathology. But
when the response is partial or moved back to a lower level, the organism finds
itself in a situation of hyper compensation response : it is obliged to deal at
a lower level with an event which would require a more general regulation. The
symptom of a pathology is going to appear as being an inadequate expression with
the tendency towards fixation, preventing the system from having a dialogue with
the world. The symptom becomes the representation of the pathology which cannot
be treated naturally : the system “displays” its pathology, each subject with
the special nature of its interpretation
(individualisation).
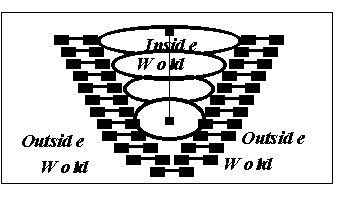
Figure
5: the self is a complex
structure
of different levels of information.
It
is the synthetic
possibility
of
moving between the various levels.
It
can have exchange functions
with
the outside word, at all levels.
The immunological self is then merged into the biological self (Figure
5): it is only its special nature which uses the immunological tools at its
disposal in order to survive. The notion of symptom, dear to homeopathy, since
it serves as a basis for the law of similarity,
can
also appear there. “Non-apparent” illnesses to which Charles Nicolle his Nobel
prize are a striking example of this. These infectious pathologies
(toxoplasmosis, rubella, etc…), due to a well-identified aggressor, may be
treated naturally by the organism without any
symptom,
thus their designation by the term “non-apparent”; however, the serological scar
is there, proving the immunological phenomenon of defence. This means that the
symptom only appears in the situation when normal behaviour cannot be exercised
(in general, during an immuno-depression, even a slight
one).
3 – The T lymphocyte receptor: semantic means of identification of the
Self
The living body does not have
the capacity of the object which is the norm of the mechanistic paradigm: it
receives and deals with information enabling it to organise its own existence.
The mechanistic interpretation of cell interactions in Immunology for instance,
results in a great difficulty in the concept of presenting the antigen to T
lymphocytes: are there any specific structures of the self which may be
identified by lymphocytes, some of which being tolerated and others rejected?
The structuralist approach is undergoing difficulties in this respect. The
immune system considered as a semantic system then comes under another
logic.
If we recall what was said in § I, antigen presenting by the competent
cells (antigen presenting cells) to T lymphocytes is always done in a pocket
made up by the terminal areas of the two chains of MHC (for class II molecules),
or by the last two domains of the MHC chain (for class I molecules). In the MHC
pocket there is a peptide able to be recognised by the receptor of a T
lymphocyte. Two situations can occur: either the peptide originates from an
“extraneous” molecule, or the peptide belongs to the self. Indeed, this pocket
is never empty and in the absence of antigenic stimulation, a peptide of the
self is going to occupy it. The lymphocyte, through its TCR receptor, is going
to identify the peptide and the MHC simultaneously (fig.
3).
Therefore,
two situations are possible (fig. 6):
i) the lymphocyte is going to identify the peptide originating from the
molecules of the self and the pair SELF/MHC + SELF/PEPTIDE will be presented to
the lymphocyte. This association does not trigger the activation of the
lymphocyte which notices no difference since in both cases it is a peptide of
the same origin, the self.
(ii) on the contrary, when the pocket is occupied by a peptide with an
extraneous origin (SELF-MHC + FOREIGN/PEPTIDE), a difference appears which
becomes significant for the immune system; indeed, from the semantic point of
view, the difference always generates a meaning: “In fact, what we call
information – the elementary information unit – is a difference which creates a
difference” (Bateson, 1980). The very fact for the extraneous peptide to be
included into the MHC molecule which is an element of the self creates the
difference. The antigen then becomes an object able to be identified by the
body: MHC makes the antigen significant to the organism. From the time this
information is identified by the appropriate T CD4 lymphocyte clone (initiators
of the immune response) which plays the role of receptor of this information, a
succession of events connected to the synthesis of cytokines and receptors
expressed on the membrane is going to provoke the clonal expansion of responding
lymphocytes which in turn are going to induce a series of cell interactions
leading to the elimination of the antigen by activation of cytotoxic cells or
through the action of antibody molecules (Bastide & Lagache, 1992, Bastide,
Lagache & Missone, 1995).

Figure
6: Meaning of the peptide
recognition
inside the MHC by the T lymphocyte
(the
peptide is included into the MHC molecule)
One
must underline that the recognition process of the antigen which triggers all the specific immunological phenomena
by analysis of “structures” (mechanistic paradigm) has not yet been elucidated.
Peptides, made up of a few amino acids, (7 to 11 for class II molecules and 10
to 17 for class I molecules), always structured in helix a,
do not make up a deciding conformational variability identifiable as a self or
“non self”. One can see that the term “non self”, classically used by
immunologists, suggests an “identification” of the antigen to be destroyed,
while the term “different from the self” only suggests the analysis of a
difference. Moreover, the positive selection of lymphocytes inside the thymus is
done only by the ability of T lymphocytes to recognise and identify the MHC,
i.e. the self.
IV –
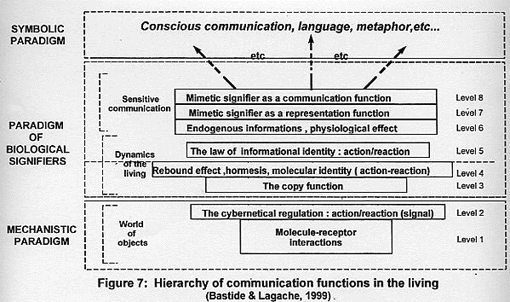
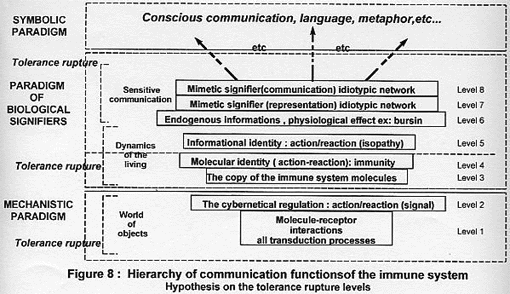
COMMUNICATION LEVELS OF THE LIVING: APPLICATION TO THE IMMUNE SYSTEM
We were able to list the various possibilities of communication in the
living (Bastide & Lagache, 1999). To do this, the use of the three paradigms
(§ III-2) is necessary to establish this hierarchy (figure 7). This brings us to
detail the paradigm of body signifiers while mechanistic and symbolic paradigms
are known in our philosophical culture.
1 – The paradigm of body signifiers
The information received by the body (§III-2) plays the role of
“biological signifiers”, able to provoke physiological alterations after
treatment of the information by the organism. This communication follows very
specific rules different from exchanges of objects (no loss after exchange but a
different situation before and after the information) (Lagache, 1988, 1997a,
1997b). The most simple examples are those which analyse experimentally the
action of endogenous substances (part of the organism itself). In the case of
highly diluted and succussed endogenous substances, therefore in an
informational form, the information provided is automatically read by the system
which “knows” its own constituents. The molecules composing the organism have a
natural meaning for this system. Thus, high succussed dilutions of bursin (a
tripeptide isolated from the Fabricius bursa of chicken) were able to “replace”
the Fabricius bursa in chickens having undergone a bursectomy to such a point
that they were able to secrete and regulate their antibodies just like normal
chicken (Youbicier-Simo & coll., 1993, 1996a, 1996b,
1997).
When, on the contrary, we are dealing with exogenous molecules the
information of which cannot be identified naturally by the organism, a “reading
frame” is necessary for the organism to be able to deal with this information.
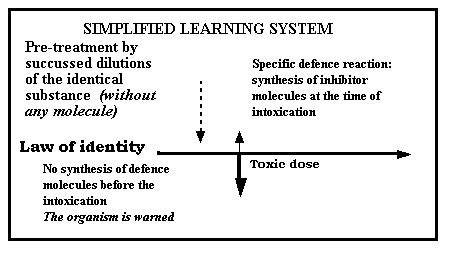
This key can be provided either by the law of identity, or by the law of
similarity suggested by homeopathy. Let us take the example of the law of identity (there is object
identity between what causes the pathology and the origin of succussed
dilutions): renal tubular cells treated with high dilutions of cadmium received
the cadmium “information” (solution concentrated at 10-40M) which is
going to provoke their resistance to intoxication by this same metal (Cal &
coll., 1986. Delbancut, 1994). In this case, we are within a law of identity: the information and the
toxic stress have the same origin and the cell was able to protect itself; the
phenomenon is strictly specific as demonstrated (Delbancut,
1994).
When we consider the law of
similarity in the homeopathic field stricto sensu, we notice that the
correspondence between the symptoms observed in the pathogenesis of a remedy
given to a healthy subject and those presented by the patient enables the
correction of the patient’s symptoms. But what is a symptom? In the patient, it
represents an expression of his
illness; an expression is nothing but the fulfilment of the self in a given
form, under the dual aegis of adaptation and totality, according to data and
history. However, the pathological circumstances have it so that the symptom is
an expression which does not result in a resolution, an uncompleted, blocked
expression. The symptom becomes then a body representation of this expression
which results from the conjunction of external and internal circumstances: it
becomes a significant creation of the individual as a whole, in his global
awareness, representing the expression the subject gives to his illness. One
finds again the Freudian notion of the symptom as healing attempt: the patient
submits to his symptoms as if they were “imposed upon” him, but they are always
an active attempt at a solution. Therefore, one understands why the homeopathic
therapy has such power: by observing the representation of the illness read in
its entirety, the therapist can reflect upon this ill body a similar image to this representation
provided by the diluted and succussed remedy. This similar image reproduces the
symptoms observed in a healthy subject having received this remedy in the
informational form. This image plays a role of re-information about the
pathology and the body is then able to treat his symptoms himself by an effect
which could be of the cathartic type (Lagache, 1988, 1997a, 1997b; Bastide &
Lagache, 1992, 1995, 1999). Therefore, the system appears as a sort of
uncompleted and somewhat misled expression which homeopathic therapy through
similarity, gets going in the direction of the capacity for change, therefore
the health balance.
Therefore, this biological signifier paradigm takes into account the
similarity, the use of highly diluted and succussed solutions and finally the
individualisation within global
awareness. It is no longer the body reduced to the condition of object of
mechanistic science, but indeed the properties of the living, its originality
and its evolution throughout time.